Tångsamhället förändras under året. De olika arterna av tång har sin huvudsakliga tillväxtperiod under vår, sommar och tidig höst. Även den associerade faunan har en tydlig säsongsvariation. Under den kalla årstiden sker mycket lite aktivitet och många av djurarterna flyttar ner till större djup, bl.a. många av fiskarterna. Många av de vanligt förekommande ryggradslösa djurarterna i tångsamhället förökar sig under våren och försommaren, t.ex. märlkräftor och havsgråsuggor. Kunskapen om säsongsdynamiken hos olika arter är dock bristfällig eftersom de flesta vetenskapliga undersökningarna sker under sommar och tidig höst.
SkagerRak- Kattegatt
Antal arter av makroalger är relativt lågt i tångbältet och förändras med årstiden. Antal arter är något högre än i Egentliga Östersjön och utöver olika grönslickar (Cladophora spp.) och tarmalgsarter (Ulva tidigare Enteromorpha spp) förekommer flera arter av fintrådiga rödalger tillhörande släktena Ceramium, Polysiphonia och Rhodomela. Några andra exempel på vanliga arter i tångbältet är rödalgerna kräkel, (Furcellaria lumbricalis) och karragenalg (Chondrus crispus). Dessa är fleråriga och förekommer under hela året medan många av de fintrådiga algerna varierar med årstiden. Typiska vår och sommararter som växer på tången är t.ex. repslick, (Spongonema tomentosum), trådslick, (Pilayella littoralis), molnslick (Ectocarpus siliculosus) och smalskägg (Dichtyosiphon foeniculaceus). Osmundea truncata liknar pepparalgen (Osmundea pinnatifida) och är vanlig som påväxt på sågtång under vår och höst. Variationen både mellan år och mellan lokaler är mycket stor.
Egentliga Östersjön
Endast ett fåtal algarter förekommer i den översta delen av algbältet och artsammansättningen förändras under året. Under våren och hösten bildar de trådformiga algerna Urospora penciliformis och gördelalg (Ulothrix spp) bälten i denna zon. Under våren dominerar vanligtvis trådslick tillsammans med grönalgerna strutsallat (Ulvopsis grevilleï) och grönborsting (Acrosiphonia spp.). På sommaren bildas ett tätt bälte av grönslick (Cladophora glomerata) med inblandning av tarmalg (Ulva tidigare Enteromorpha intestinalis) samt brunalgerna brunsnärja (Scytosiphon lomentaria) och skäggalg (Dictyosiphon foeniculaceus) och under sensommar dominerar rödsläke (Ceramium) zonen. Dessa arter kan delvis förekomma som påväxt på tången och delvis växer de direkt på klippan. Under tångplantorna är klippan ofta täckt av havsstenhinna (Hildenbrandia rubra).
Bottenhavet
Blåstång och smaltång är fleråriga arter som förökar sig under sommarhalvåret. Säsongsvariationen hos de ettåriga alger som förekommer i tångbältet liknar i hög grad den i Egentliga Östersjön, men produktionssäsongen är ofta kortare och vårsäsongen förlängd. De vanligaste arterna är trådslick (Pilayella) och ullsläke (Ceramium tenuicorne), som förekommer under alla tider på året. Närmast ytan ses ofta en produktionstopp av grönslick (Cladophora arter) under sommaren, och ullsläke under hösten. Efter milda vintrar finns ullsläke kvar på klipporna nära ytan.
Förändringar av tångsamhället i Östersjön sedan 1940-talet.
Under de senaste decennierna har tidigare opåverkade kustområden i Östersjön visat tydliga tecken på påverkan av både lokal och storskalig övergödning. Den mest markerade effekten av övergödningen har varit en storskalig tillbakagång av fleråriga makrofyter/alger som t.ex. blåstång och sågtång, både areellt och i fråga om minskad djuputbredning. Till exempel har djuputbredning av blåstång minskat med flera meter mellan 1944/45 och 1984 [31]. Orsaken till dessa förändringar är en respons på en ökad grumlighet beroende på planktonblomningar och skuggning av epifyter, ökad sedimentering av organiskt material samt i vissa områden betning av havsgråsuggor (Idotea baltica). Det finns en sammanställning av utbredning och förändringar i den bentiska makroalgvegetationen i Dahlgren och Kautsky [40].
När närsaltshalterna ökar gynnas snabbväxande fintrådiga och bladlika alger som grönslick (Cladophora glomerata), havssallat (Ulva lactuca), strutsallat (Monostroma grevillei) och tarmalg (Ulva intestinalis) . På de hårda bottnarna ersätts tångbältet av fintrådiga röd- och brunalger som t.ex. trådslick (Pilayella littoralis), molnslick (Ectocarpus siliculosus), ullsläke (Ceramium tenuicorne), rödris (Rhodomela confervoides)och fjäderslick (Polysiphonia fucoides) eller av tunna bladlika alger som t.ex. rödalgen ekbladsalg (Phycodrys rubens). Dessa kortlivade arter, lossnar från botten mot slutet av växtsäsongen och efter stormar. De lösa algerna ansamlas och bildar drivande algmattor på större djup [41] eller driver in till kusten och sköljs upp på land [5].
Sedan 1970-talet har avloppsvatten renats på i första hand fosfor och senare även kväve, vilket lett till minskade halter i vattnet t.ex.
i de inre delarna av Stockholms skärgård (se vidare Svealands kustvattenvårdsförbund).
Någon tydlig återhämtning
av blåstångens förekomst i de innersta delarna har dock inte noterats [42]. Det enda område utmed den svenska kusten där det finns
information om hur tångbältets djuputbredning och biomassa förändrats sedan 1970-talet är i Asköområdet, beläget
i Sörmlands skärgård [43], [29], [30]. Där har genom det nationella miljöövervakningsprogrammet ett större antal provtagningslokaler
följts årligen sedan 1993. Här minskade djuputbredningen av makroalgsvegetationen med 0,7 m från 10,6 meter 1974/75 till 9,9 meter till
1994. På motsvarade sätt minskade blåstångens djuputbredning med 1-2 m mellan 1974/75 till 1994 för att sedan under den senast
10-årsperioden vara mer eller mindre oförändrad. Medeldjupet för blåstången i området är nu runt 4 meter både
i inner- och ytterskärgården. En art som förekommer i tångbältet men även på djupare bottnar är den fleråriga
rödalgen rödris (Rhodomela confervoides), vilken har minskat under hela 1990-talet fram till nu [30]. Orsaken till denna minskning är
inte klarlagd men en trolig förklaring kan vara en fortsatt hög sedimentation av partiklar på lite djupare bottnar. Hur stora förlusterna är
i täckningsgrad och biomassa varierar mellan olika områden. Utmed Blekinge kusten minskade förekomsten av blåstång och sågtång
under 1990-talet [44]. Samtidigt förändrades den relativa dominansen från blåstång till sågtång. Minskningen av blåstång
skedde samtidigt som stora mängder med havsgråsuggor förekom i tångbältet och dessa föredrog att beta av blåstång
speciellt vid högre vattenomsättning och vågexponering [44].
En av de få studier av direkta förändringar i artantal i samband med övergödning är ifrån Ålands hav, där man
funnit att sex arter försvann mellan 1956 och1993 och att det artrikaste algsamhället påträffades redan på 4-7 m djup år 1993,
jämfört med 10-15 m djup år 1956 [45]. Detta innebär att ett antal arter troligen blivit mer ovanliga/alternativt helt försvunnit
och att den artrikedom som förekommer nu finns grundare än tidigare. Det är även möjligt att tidiga vårarter kan ha försvunnit
eller kommer att försvinna eller minska i utbredning utan att detta upptäcks, eftersom ytterst få studier görs under våren och försommaren.
Tångbältets genetiska biodiversitet
Tångens genetiska variation
Genetisk variation hos blåstång (Fucus vesiculosus).
Blåstång är en mycket variabel art, både när det gäller form, fysiologiska anpassningar och tidpunkt för förökning. Kartläggning av DNA-sekvenser hos populationer av blåstång på ost- och västkusten visar också en stor genetisk variation. Genetiska skillnader mellan plantor som växer på olika lokaler förekommer generellt på avstånd ned till 1 km, och i vissa fall ned till så korta avstånd som 10 m [46]. Stora genetiska skillnader finns mellan plantor som växer på svenska ostkusten och de som växer på svenska västkusten. Jämför man nivån på denna skillnad med motsvarande nivå hos t.ex. torsk (vars Östersjö- och Kattegatt/Skagerrak bestånd också skiljer sig genetiskt från varandra) så är blåstångens skillnader cirka 3-4 ggr större.
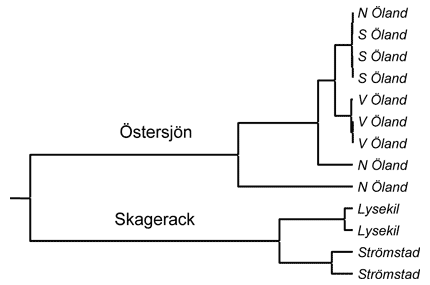
Populationer av blåstång ifrån Östersjön och Skagerack visar tydliga genetiska skillnader i neutrala anlag (anlag som inte påverkar överlevnaden). Dessa skillnader är främst orsakade av ett mycket begränsat utbyte av gener mellan de båda haven.
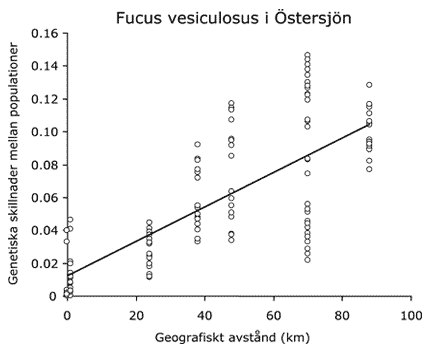
För blåstång ökar den genetiska skillnaden mellan bestånd snabbt med ökande geografiskt avstånd mellan bestånden.
De stora genetiska skillnaderna mellan individer av blåstång på olika lokaler är en konsekvens av att förökningskropparna (de befruktade äggen) har en begränsad spridningskapacitet och hamnar helt nära moderplantan. Eftersom sommar- och höstförökande blåstång har olika tider för reproduktion [47] så skulle man kunna förvänta sig att finna genetiska skillnader även mellan dessa grupper, men så är inte fallet. Mellan bestånd av sommar- och höstförökande blåstång finns inte större genetiska skillnader än vad man förväntar sig mellan bestånd på motsvarande avstånd som förökar sig vid samma tid [46]. Inga undersökningar är ännu gjorda på eventuella skillnader mellan individer av de två typerna där de fimms i blandade bestånd.
Blåstång förökar sig sexuellt i de flesta miljöer. Hanplantor släpper ut spermier och honplantor ägg fritt i vattnet (i samband med fullmåne och nymåne), om det är lugnt i vattenmassan [48]. Blåstång i Östersjön, och speciellt i norra Östersjön, uppvisar överraskande nog en tendens till kloning (se F. radicans nedan). Blåstång kan hybridisera med andra arter inom släktet Fucus.
Eftersom blåstången består av lokalt avgränsade genetiskt olika bestånd är det troligt att även egenskaper som är betydelsefulla för blåstångens överlevnad och reproduktiva framgång förmodligen är starkt påverkade av lokala förhållanden. Detta innebär att lokala bestånd kan utveckla anpassningar till den miljö de lever i. Genom att transplantera plantor från Östersjön till västkusten har det gått att påvisa att Östersjöns blåstångsindivider saknar egenskaper som är av stor betydelse för att kunna överleva på västkusten, t.ex. tål de inte infrysning vilket västkustplantor gör [17].Genetisk variation hos smaltång (Fucus radicans)
Från Åland och norrut finns en liten tångart som är smalare än blåstång och som saknar flytblåsor. Man har tidigare ansett att denna smalbladiga tång är en liten variant av blåstång, men genetiska studier visar att den är en egen art, som döpts till smaltång (Fucus radicans) [6]. För närvarande är Bottenhavet enda kända växtplatsen för denna art, men det kan inte uteslutas att den även finns i andra områden, förmodligen nordligt (Grönland, Vita Havet). Smaltång lever ofta i blandade bestånd med småväxt blåstång som också är allmän upp till Höga Kusten.
De två populationer av smaltång som hittills har analyserats genetiskt (Järnäs i norra Bottenhavet och Öregrund, Upplands skärgård) uppvisar en ovanlig egenskap; de flesta individerna (>80%) har uppkommit genom kloning [49]. Kloning av fastsittande plantor har aldrig tidigare beskrivits inom gruppen stora brunalger (blåstång, tare och dess släktingar). Intressant nog uppvisar även Östersjöbestånden av blåstång (F. vesiculosus) kloning, ~30% av individerna är klonade vid Öregrund och <10% på Öland. Kloning leder till kraftigt minskad genetisk variation inom och mellan populationer. Hos smaltång i Öregrund och Järnäs har ca 70-80 % av individerna exakt samma uppsättning av genvarianter. Om denna klon skulle slås ut (t.ex. av ett extremt miljötillstånd) kommer alltså 70-80% av bestånden att försvinna.Genetisk variation hos sågtång (Fucus serratus)
Graden av genetisk variation hos sågtång liknar i stor utsträckning den hos blåstång. På avstånd av någon kilometer är grupper av individer i stort sett alltid genetiskt något olika [50]. På samma sätt som för blåstång är förmodligen lokala bestånd genetiskt anpassade till sin lokala miljö, och det genetiska utbytet mellan bestånd minskar kraftigt med ökande geografiskt avstånd. Sågtången förökar sig sexuellt och huvuddelen av bestånden är fertila på hösten och senvintern, men en population av sommarförökande sågtång har noterats vid sydkusten på Öland [51]. Reproduktion genom kloning är inte känt hos sågtång.
Andra makroalgsarter
Idag finns information om genetisk variation och struktur hos svenska populationer av ytterligare några arter av marina alger: Ceramium (ullsläke) är mångformig i sitt utbredningsområde och ansågs tidigare bestå av tre arter C. tenuicorne, C. gobii och C. strictum. Genetiska studier har dock visat att de är samma art, dvs. att det finns ett genflöde mellan representanter för de olika grupperna och korsningsförsök visar att de även kan föröka sig sinsemellan [52]. Enligt ekofysiologiska studier kvarstår dock tecken på att olika populationer är lokalt anpassade, t.ex. ullsläke från inre Östersjön tål låg salthalt bättre än ullsläke från Egentliga Östersjön [53].Genetisk variation hos ryggradslösa djur i tångbältet
För vissa djurgrupper i tångbältet finns idag jämförelsevis stor genetisk kunskap. En av dessa grupper är strandsnäckor (Littorina) där tre av Sveriges fyra arter lever i den övre delen av tångbältet (trubbig strandsnäcka Littorina fabalis, L. obtusata och vanlig strandsnäcka L. littorea ) och den fjärde arten, levandefödande strandsnäcka (L. saxatilis) lever på klipporna strax ovanför tången. Dessa fyra arter exemplifierar två livsstilar som är vanliga för marina bottenlevande ryggradslösa djur: (1) arter med ett långt planktonlevande larvstadium och (2) arter som saknar planktonlevande larv eller endast har ett kort planktonlevande stadium. Att ha planktonlevande larv eller inte påverkar starkt den genetiska strukturen hos strandsnäckorna. Andra studier visar att ungefär samma mönster som gäller strandsnäckorna också gäller andra arter av ryggradslösa djur, som i sitt vuxna stadium är fastsittande eller mindre rörliga. Mönstret är följande: Marina arter med ingen eller dålig larvspridning är kraftigt genetiskt strukturerade, dvs. man finner genetiska skillnader mellan populationer av individer på korta avstånd (<1-10 km). Marina arter med bra larvspridning är oftast svagt eller inte alls genetiskt strukturerade över ganska stora avstånd (>10-100 km).
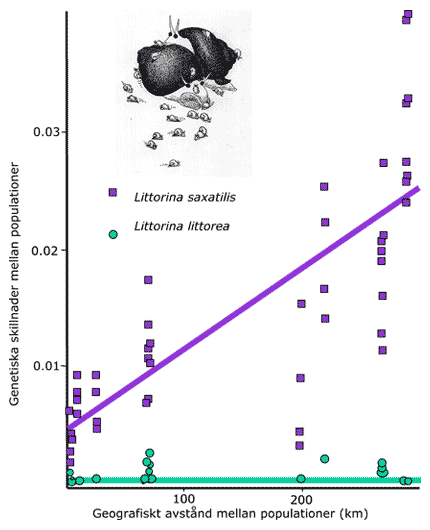
Genetiska skillnader mellan individer/populationer hos en art som saknar larver (här levande födande strandsnäcka, Littorina saxatilis) ökar snabbt med ökande geografiskt avstånd. Hos en art med larvspridning (här vanlig strandsnäcka L. littorea) uteblir genetiska skillnader över samma geografiska avstånd [54].
De genetiska skillnaderna som visas i figuren ovan är skillnader uppmätta på molekylär nivå (proteiner/DNA). På motsvarande sätt finns stora skillnader i storlek, form och funktion mellan populationer hos levandefödande strandsnäcka, medan sådana skillnader nästan saknas helt hos vanlig strandsnäcka. Levandefödande strandsnäcka är i hög grad lokalt anpassad till sin livsmiljö, medan arten vanlig strandsnäcka har generella egenskaper. Detta betyder att för att kunna skydda eller genomföra en restaureringsåtgärd så måste man ta hänsyn till om arten ifråga har en hög lokal anpassning eller ej.
Tabellen nedan beskriver hur ekologiskt viktiga egenskaper varierar mellan miljöer för den arten av strandsnäcka som saknar larvspridning och är kraftigt genetiskt strukturerad. För arten med larvspridning och vag genetisk struktur saknas variation. Precis som för strandsnäckorna har studier på andra arter visat att det oftast finns ett samband mellan om en art har en god spridning eller ej och i vilken utsträckning arten är genetisk strukturerad över kortare geografiska skalor. Det finns dock viktiga undantag från detta samband och viktiga sådana är de markanta genetiska förändringar som sker i ett stort antal arter (även arter med god spridningsförmåga) i och runt Öresund, i skiftet mellan Egentliga Östersjön och Västerhavet, se vidare under Genetisk mångfald/Marin genetisk mångfald/Figur 3.
Art |
Littorina littorea |
Littorina saxatilis |
|---|---|---|
Svenskt namn |
Vanlig strandsnäcka |
Levandefödande strandsnäcka |
Larvutveckling |
Planktoniska larver |
Saknar fria larver |
Genetisk struktur |
Inga genetiska skillnader utmed svenska kusten |
Genetiska skillnader går att mäta redan från avstånd på 10-tals meter |
Lokala anpassningar |
Inga ärftliga lokala anpassningar |
Stora ärftliga lokala anpassningar: - |
|
(viss utseendevariation mellan miljöer orsakas av direkt miljöpåverkan) |
storlek, form, skalets tjocklek, skalets färg, beteende, tålighet mot sötvatten och torka, mm. |
Reproduktionshinder inom arten |
Inget känt (ej särskilt troligt) |
Reproduktionshinder mellan individer från olika miljöer pga dess olika utseende och egenskaper |
Grad av genetisk biodiversitet i gener som påverkar individens egenskaper |
Låg genetisk biodiversitet inom arten |
Hög genetisk biodiversitet inom arten |
Men inom Västerhavets tångbälten och inom Östersjöns tångbälten kan man utifrån en arts spridningsförmåga någotsånär förutsäga om arten har en stor eller en mindre variabel genetisk struktur. Några vanliga grupper av tångbältesorganismer listas i tabellen här nedanför. Tabellen anger också säkerheten i bedömning, där färgkoden visar om det finns genetiska belägg från många arter (hög säkerhet), några arter (måttlig säkerhet) eller inga arter (låg säkerhet, bedömning gjord med utgångspunkt i spridningspotentialen).
Gruppens dominerande reproduktionsstrategi |
Har pelagiskt larvstadium |
Saknar pelagiskt |
|---|---|---|
Förmodad grad av lokal anpassning |
Låg |
Hög |
Havsgråsuggor |
|
X |
Märlkräftor |
|
X |
Havstulpaner |
X |
|
Tångräkor/krabbor |
X |
|
Sjöstjärnor och sjöborrar |
X |
|
Snäckor |
X |
X* |
Musslor |
X |
|
Hydroider |
X |
|
Mossdjur |
X |
|
Havsborstmaskar |
X |
X** |
*De flesta arter har larv. Undantag bland vanliga arter i tångbältet är: Littorina obtusata (trubbig strandsnäcka), Littorina fabalis (trubbig strandsnäcka), Nucella lapillus (purpursnäcka), tusensnäckor Hydrobia ventrosa (tusensnäcka),
**Vissa arter saknar larv, t.ex. Hediste (Nereis diversicolor)
